No evidence for niche competition in the extinction of the South American saber-tooth species

Introduction
The South American continent remained in isolation for a long time over the Cenozoic, resulting in a peculiar mammal assemblage structure, such as the endemic metatherian apex predators1,2. However, the Great American Biotic Interchange (GABI) ended the South American isolation and promoted the arrival of the North American carnivores3. The GABI changed expressively the history of mammals in South America, where almost half of its current fauna has North American origin4. Even so, it is still unknown how this fauna interacted when they came into connection.
The contact between faunas that evolved in isolation was already pointed out to be responsible for South American mammalian extinctions5. The order Carnivora (Eutheria) arrived in South America between 9 and 3 Ma (pre-GABI, see6), the time at which the extinction rate of Sparassodonta (Methateria) increased7. Since carnivores and sparassodonts are typical examples of evolutionary convergence (e.g., both have fox-like and saber-toothed forms2,8), it has been proposed that Sparassodonta became extinct during the Plio-Pleistocene due to competition with placental Carnivora from North America4,9, a classic example of the competitive exclusion principle, by which species with identical niches cannot coexist indefinitely10. This principle is based on the fact that resources are limited to some extent, and species that exploit similar resources naturally compete for them11. Under such a scenario, the weaker competitor perishes, and the more skilled one survives.
Recent studies have pointed out that, although placental carnivores replaced the Sparassodonta in South America, there is no clear evidence of competition between them7,12,13. We could infer that competition was the cause of extinction if the species have indeed co-occurred in time and space, and played a similar ecological function, which seems to be the case for South American carnivores and sparassodonts. Patterns of species co-occurrence in space and time can be investigated from the perspective of climatic niche overlap or differentiation, however, this has not been done hitherto. On the other hand, the extinction of Sparassodonta may have been caused by other events, such as the extinction of their prey and environmental changes related to the Andean uplift, atmospheric CO², temperature, and sea level7,13.
Here, we used paleoclimatic data to compare the climatic niche of two saber-toothed groups, Sparassodonta, represented by the species Thylacosmilus atrox (Riggs, 1933), and Carnivora, represented by two species of Smilodon (Lund, 1842): S. fatalis (Leidy, 1868) and S. populator (Lund, 1842). These two groups of large-sized predators inhabited open vegetation areas of South America ~6 Ma4 and preyed upon large herbivores2,12,14,15,16. We evaluated both groups’ niche width, overlap, and similarity. If there is climatic niche overlap between T. atrox and Smilodon, these groups could have co-occurred, and the competitive exclusion of Sparassodonta cannot be discarded. We also expect higher climatic niche overlap between T. atrox and S. populator because S. populator was more widely distributed in South America in comparison with S. fatalis17. On the other hand, the absence of niche overlap and niche similarity makes way for other causes (such as environmental changes) for Sparassodonta extinction.
Results
Climatic niche width, similarity, and overlap
Thylacosmilus atrox presented a narrower climatic niche width in comparison to S. fatalis and S. populator, except for the mean temperature of the warmest quarter, which is wider in T. atrox. Smilodon fatalis presented a narrower climatic niche in comparison to S. populator, except for the mean temperature of coldest quarter (Fig. 1). Proportionally, the climatic niche width of T. atrox vary from 0.31–0.6 in comparison to S. fatalis and S. populator, except for mean temperature of warmest quarter, which was wider in T. atrox than in S. fatalis and corresponds to 0.71 – S. populator’s niche width (Table 1).

Climatic variation from the occurrence fossil records of South and North American saber-tooth predators.
The results of niche overlap and similarity were quite similar in both time intervals (5 Ma and between 3.8 and 1.8 Ma). Thus, hereafter we only kept the results for the time interval between 3.8 and 1.8 Ma (the results for the whole period of 5 Ma are provided in Supplementary information). The similarity of the climatic niche between T. atrox and S. fatalis was higher than expected from random (p = 0.005 for Schoener’s D, and p < 0.005 for Hellinger’s I), but the similarity between T. atrox and S. populator did not differ from random expectations (p = 0.14 for Schoener’s D, and p = 0.15 for Hellinger’s I). Additionally, there was considerable niche overlap between T. atrox and S. fatalis (Schoener’s D = 0.34 and Hellinger’s I = 0.52 Fig. 2a), and low niche overlap between T. atrox and S. populator (Schoener’s D = 0.1 and Hellinger’s I = 0.14, Fig. 2b).
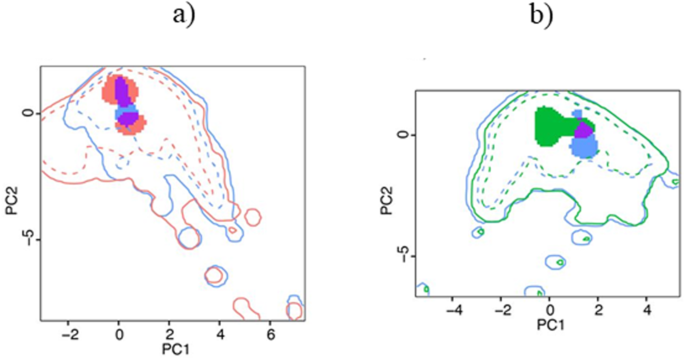
Niche overlap between T. atrox (in blue) and S. fatalis (in red) (a), and between T. atrox and S. populator (in green) (b). Purple color indicates overlapping between two species. The solid and dashed lines represent 100% and 50% of the available climatic space, respectively.
Discussion
We found no evidence of competition between T. atrox and S. populator, suggesting they occurred in different climatic conditions and, probably, did not overlap geographically. On the other hand, although the niche similarity and overlap between T. atrox and S. fatalis could indicate a competition signal, these species probably also did not co-occur (the distribution of T. atrox was restricted to Argentina, where no record for S. fatalis is known, Fig. 3a17,18). Rather, the niche similarity between them could be due to the broader climatic niche of S. fatalis, which could comprise climatic conditions also suitable for T. atrox, although in distinct geographic areas.
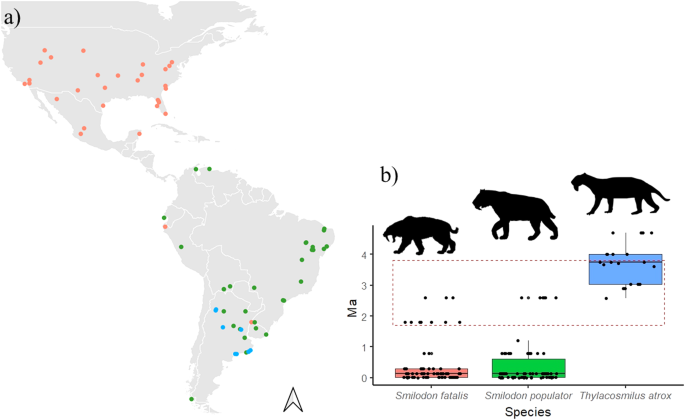
a Spatial and b temporal (in millions of years—Ma) distribution of the occurrence records of Thylacosmilus atrox, Smilodon fatalis, and Smilodon populator in the American continent. The red dashed box indicates the time slice of 2 Ma (from 3.8 to 1.8 Ma) for which niche overlap and similarity were estimated. The silhouettes were obtained from PhyloPic https://www.phylopic.org/. Thylacosmilus atrox was designed by Zamices (Julián Bayona), under the Creative Commons Attribution-NonComercial 3.0 Unported license https://creativecommons.org/licenses/by-nc/3.0/ no changes were made. Smilodon fatalis was designed by Steven Traver, under the Creative Commons CC0 1.0 Universal Public Domain Dedication license (https://creativecommons.org/publicdomain/zero/1.0/) no changes were made. Smilodon (considered here as S. populator for illustration purposes) was designed by Margot Michaud under Creative Commons CC0 1.0 Universal Public Domain Dedication license (https://creativecommons.org/publicdomain/zero/1.0/) no changes were made.
We found that the South American T. atrox had a narrower climatic niche compared to the North American Smilodon species. However, the interpretation of niche width for T. atrox should be cautious due to the lack of information about the distribution of this species (n = 20 occurrence records), which may have contributed to the narrower niche estimate found here. Despite this, our results corroborate previous works16, which found a narrower niche diet for T. atrox in comparison with Smilodon. Species with narrower climatic niche width may have lower tolerance to climate change, and narrower temperature niche width has been associated with species extinction19. Also, narrower thermal tolerance has already been related to climatic change vulnerability in mountain species with low dispersion capacity, and in species with long generation times20,21 (e.g., large-sized mammals). Therefore, the more restrictive distribution of T. atrox in the southern portion of South America (Fig. 3), and its narrower climatic niche together with its larger body size (117.36 kg2,15) suggest that this species could have been more vulnerable to environmental changes than its potential competitors Smilodon species.
Our results corroborate previous results that failed to find competition evidence between Sparassodonta and Carnivora2,7,12,13,22. Thus, other factors still unknown may have been responsible for the South American apex predator extinction. Indeed, recent studies agreed that high extinction rates in Sparassodonta started in the Miocene (16 Ma13, 17 Ma7) and remained until the group extinction in Pliocene13. Changes in regional landscape due to Andean uplift, atmospheric CO², and sea level were pointed as the cause for Sparassodonta extinction7,13. Here, we provide additional support for these causes, showing that T. atrox had a narrower climatic niche than the other species investigated herein and, therefore, could have been more vulnerable to climatic change20,21.
It should be highlighted that several caveats may affect the interpretation of our results. We are dealing with fossil information for the last 5 Ma, with inaccurate dates and clear distribution gaps throughout the study area (Fig. 3a). Furthermore, the narrowed time slice from 3.8 to 1.8 Ma reduced the number of occurrence records available for niche overlap analyses, precluding more thinned analyses (e.g., glaciation and interglaciation fluctuations). Also, the coarse resolution (1 degree of resolution20) of climate data available for that period has limitations for investigating species interaction locally. Despite this, the analysis including all available fossil records showed similar results, thus providing additional support to the patterns unveiled in this study. Indeed, our results add up with previous findings of no competitive interaction between sparassodonts and carnivores, thus increasing the body of evidence for the potential causes of South American saber-tooth extinction.
Methods
Occurrence and paleoclimatic data
We obtained fossil occurrence data of T. atrox, S. fatalis, and S. populator spanning from 5 Ma to the present, the period for which the climate data was available (see below), from The Paleobiology Database (PBDB https://paleobiodb.org). Occurrence records of T. atrox were also obtained from literature7 due to the low number of occurrences available in the PBDB. We retrieved 156 occurrence records (20 for T. atrox7,23, 70 for S. fatalis and 66 for S. populator24, Fig. 3). We extracted climate values for each fossil occurrence for the only four bioclimatic variables available in the PALEO-PGEM-Series25 (1-degree of resolution): mean temperature of warmest quarter, mean temperature of coldest quarter, mean precipitation of wettest quarter, and mean precipitation of driest quarter. We used the “raster” package26 for extracting climate data.
Climatic niche width, similarity, and overlap
To compare the climatic niche of T. atrox and Smilodon species, we calculated the niche width, i.e., the difference between the highest and the lowest values for each bioclimatic variable for each species. Then, we calculated the proportion of the T. atrox niche width concerning the Smilodon species, dividing the width value of T. atrox by the width value of both Smilodon species.
The temporal resolution of 5 Ma can be considered too coarse to interpret potential changes in species niche due to the magnitude of climate variability during the Plio-Pleistocene. Thus, for niche overlap and similarity analysis, we narrowed the time slice to the period between 3.8 and 1.8 Ma (Fig. 3b, red dashed box), which encompasses a period of higher overlap among the records for all species analyzed here. Within this period of 2 Ma we keep 12 fossil records for T. atrox, 10 for S. fatalis, and 6 for S. populator (Fig. 3b). We also estimated the niche metrics based the whole period of 5 Ma to assess the consistency of the results through different time intervals (i.e., 2 Ma and 5 Ma). In this case, we applied a bootstrap approach to account for the differences in the number of fossil records among the species (from 20 for T. atrox to 70 for S. fatalis; see Supplementary information).
We conducted the niche overlap analyses using “ecospat” package27,28,29,30, on R environment31. The distribution of T. atrox and S. populator fossil records was restricted to South America, while S. fatalis had fossil records throughout the whole American continent (Fig. 3). For both regions (South America and the whole American continent) we created two environmental grids, one composed by the fossil records and the other by the background area. Therefore, the background area for T. atrox and S. populator was South America, while for S. fatalis the background area was the American continent. We obtained background areas for each year of the occurrence records for each species.
We performed a Principal Component Analysis (PCA) using the “dudi.pca” function of the ade4 package32 using the environmental grids, one PCA for each background area. Subsequently, we used the two first PCA axes to create a kernel density estimation using the “ecospat.grid.clim.dyn” function. We then tested the similarity between the niches using the “ecospat.niche.similarity.test” function, from which we extracted the probability of the similarity being higher than randomly, for Schoener’s D and Hellinger’s I indices. We also calculated the Schoener’s D and Hellinger’s I indices to assess species’ niche overlap. Finally, we plot the niches in environmental space using “ecospat.plot.niche.dyn”. These analyses were performed separately for each species interaction: between T. atrox and S. fatalis, and between T. atrox and S. populator.

Responses